- КАРИО-
КАРИО- (от греч. karyon—ядро): кариогамия—слияние ядер при половом процессе; кариокинез—деление ядра с образованием хромосом (кариомитоз); кариолимфа— «ядерный сок»; кариолизис—растворение ядра при дегенерации; кариомеры—зернистые участки, из к-рых состоят хромосомы на стадии спиремы во время кариокинеза; карио-мериты—ядра, образовавшиеся из материнского ядра, но содержащие лишь часть нормального числа хромосом или даже одну хромосому; кариоплазма—ядерное вещество в отличие от вещества тела клетки протоплазмы; кариосома—т.н.ядрышко, или ну-клеолус ядер высших животных и растений и нек-рых простейших. НАРИОНИНЕЗ, karyokinesis (Schleicher) (от греч. karyon—ядро и kinesis—движение), син. митоз (Flemming), непрямое деление ядра (indirekte Kernteilung; Flemming), сложный процесс клеточного деления, являющийся нормальным способом размножения клеток в противоположность амитозу (см.), или прямому делению. История. Размножение клеток путем деления было установлено в 40-х и 50-х гг. 19 века Негели (Naegeli) для растений и Ремаком (Remak) для животных. Оно описывалось просто: деление ядрышка, перешнурование ядра на 2 части с последующим разделением клеточного тела. В 70-х гг. в связи с усовершенствованием микроскопа (иммерсия) и методов исследования оказалось, что деление I клетки происходит много сложнее. Первое подробное описание деления было дано для растительных клетокШтрасбургером (Strass-burger; 1875) и русским ботаником Чистяковым; для животных—в 1878 г. Шлейхером (Sehleicher; хрящевые клетки), киевским профессором Перемежко (личинка тритона) и гл. обр. Флеммингом (личинка саламандры). Последний в своих классических работах установил главные фазы К. и выработал методику, применяемую и до наст, времени. В 80-х и 90-х гг. трудами целого ряда выдающихся исследователей (Balbiani, van Beneden, Henneguy, Carnoy, С. Rabl, Hermann, Druner, Boveri, Kostanecki, M. Hei-denhain). В схему Флемминга были внесены значительные дополнения; было доказано повсеместное распространение К. и изучен ряд важных закономерно повторяющихся отклонений от общей схемы. В 90-х гг. стали появляться попытки объяснения механизма К. В виду того, что изучение фиксированных препаратов не дало возможности разре-,шить многие связанные с К. проблемы, за последнее десятилетие возобновилось прижизненное изучение кариокинеза (преимущественно в тканевых культурах), дополняемое применением микродиссекции (Chambers, Speck, Belar). С х е м а К. Изменения клетки при К. слагаются 1) из тех изменений, к-рые наблюдаются в ядре, гл. обр. его хроматине, и ведут к образованию хроматиновых отдельно-стей, хромосом, к-рые затем расщепляются по длине и распределяются поровну между дочерними клетками (хроматиновая фигура); 2) из изменений в плазме: деления центросом, возникновения веретена и лучистых сияний, играющих роль в процессе деления клеточного тела и распределения хромосом (ахроматиновая фигура). Оба эти процесса могут до известной степени протекать самостоятельно, но при нормальном К. строго координированы. К. в животных клетках в общем протекает одинаково; в основу схемы, обычно излагаемой в учебниках, кладутся

10 11 12
Рисунок 1. Кариокинез у тритона; хроматиновая фигупа: I—покоящееся ядро; 2 и 3—стадий клубка; 4, 5 и 6—стадий материнской звезды; 7 и «—стадий метакинеза; 9 и 10—стадий дочерних звезд; 11—стадий дочерних к уб^ов; 12—покоящиеся ядра. (По Пете^сену.) картины, наблюдаемые у хвостатых амфибий (рис. 1) и при делении яйца лошадиной аскариды (для ахроматиновой фиг.) (рис. 2).— Процесс К. принято разделять на 4 главные фазы: про-, мета-, ана- и телофазу (Strassbur-ger,M. Heidenhain)mm на ряд последовательных фигур (Flemming). Профаза начинается изменением ядра, которое набухает и округляется.Количество хроматиновых зерен 1 2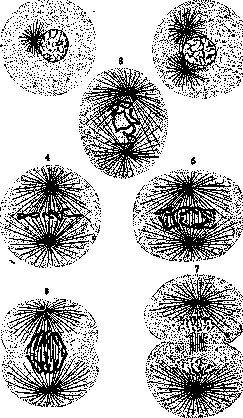
Рисунок 2. Полусхематическое изображение кариокинеза у Ascaris megalocephala: i—покоящаяся клетка; 2—деление центросомы; з— профаза: центросомы располагаются по полюсам, лучи сильно развиты; хроматин распался на хромосомы; 4—материнская звезда; хромосомы расположены по экватору; 5—метафа-за: хромососы, расщепившиеся по длине, удаляются к полюсам; в—анафаза—начинающееся деление клеточного тела; 7—деление клеточного тела почти закончилось; ядра переходят в стадий клубка. (По Кастенецкому.)
в нем постепенно нарастает; сближаясь между собой, они начинают собираться в нити, и в ядре образуется хроматиновая фигура, получившая название клубка (с п и ре м а), в виде длинной, чрезвычайно извитой нити, располагающейся преимущественно периферически. Впоследствии нить становится короче и толще, клубок как бы разрыхляется, вследствие чего принято различать два стадия: плотного клубка (ранняя профаза) и рыхлого клубка (поздняя профаза). Клубок распадается далее на отдельные участки, сегменты,—хромосомы (Waldeyer). Необходимо заметить однако, что последние очень часто выступают сразу в виде отделыгостей. Одновременно с изменением ядра замечаются изменения в протоплазме: центросома, к-рая в покоящейся клетке видна далеко не всегда, выступает
Рисунок 3. Деление центросомы; зерна Пфи-цнера; раннее расщепление
хромосом. (По Герману.) отчетливо в виде маленького пузырька, окружается лучистым сиянием, затем делится, и между отступающими половинками ее протягивается волокнистое образование, имеющее вид веретена (рис. 3). Совокупность веретена, центросом по его концам и лучистых сияний (полярные сияния, астеры, аст-росферы), идущих от них, вследствие слабой окрашиваемости при обычных гист. окрасках получила название ахроматиновой фигуры (иначе—амфиастер). Однажды образовавшись, она продолжает расти в течение всей профазы и вступает в связь с хромосомами.Происходит это таким образом, что в известный момент ядро утрачивает свою оболочку (она растворяется), ядрышки также или растворяются или распадаются на мелкие обломки, и на месте ядра получается светлая ядерная полость (Kernhohle), в которой располагаются хромосомы. Тогда между ними и центросомами становятся заметными тонкие соединительные волоконца, вырастающие от центросом и связывающие каждую хромосому с обоими центрами (тянущие волокна, «вожжи»; рис. 4). Веретено изменяет свое место, поворачивается и устанавливается посередине клетки, в ядерной полости, а хромосомы располагаются по его экватору в равном расстоянии от полюсов— экваториальная пластинка, или материнская звезда (м о н а с т е р). С установкой веретена К. вступает в мета фазу,в течение к-рой хромосомы материнского ядра расщепляются по длине на тождественные друг с другом дочерние хромосомы и начинают постепенно отходить друг от друга, оставаясь нек-рое время в соединении концами (перераспределение, или метакинез). Эта фаза без резких границ переходит в анафазу, когда хромосомы, окончательно отделившись друг от друга, продолжают двигаться к полюсам, скользя по поверхности веретена (как предполагали прежде, вследствие сокращения идущих к ним волоконец); дойдя до полюсов, они останавливаются, образуя фигуру дочерних звезд (д и а -стер). В это время начинается разделение клеточного тела: полярные сияния сильно вырастают, доходя почти до периферии, и в месте их перекрещивания, по экватору клетки, намечается круговая борозда— перешнурование. Конечная фаза, т е л о ф а -з а, характеризуется тем, что дочерние ядра переходят в покоящееся состояние, проделывая в обратном направлении изменения профазы (реконструкция ядра). Хромосомы дочерних звезд сближаются, иногда соединяются концами, образуя фигуры дочерних клубков (д и с п и р е м а); по периферии их появляется тонкая ядерная оболочка. Она сначала плотно облегает концы хромосом, затем
Рисунок 4. Образование ахро-
матинового веретена (по Герману). по мере накопления ядерного сока отстает от них, и ядро округляется. Хромосомы теряют гладкие контуры, покрываются шишками, соединяются друг с другом мостиками, и хроматин распределяется по образовавшейся сети в виде глыбок и зерен. С появлением оболочки в ядре вновь появляются ядрышки—одно или несколько. В то же время деление тела продолжается, и круговая борозда окончательно отделяет дочерние клетки, к-рые остаются известное время в соединении только остатками веретена в виде узкого мостика. Полярные сияния и веретено становятся постепенно все менее резкими и постепенно исчезают, образуя иногда зернистое скопление (архоплазма), а центросомы переходят от полярной части ядра на другую сторону (телокинез); иногда это передвижение сопровождается вращением ядра (Гейденгайн). Такова картина К. на фиксированных препаратах; экспериментальные прижизненные наблюдения последних лет в существенных чертах ее подтверждают, внося нек-рые изменения лишь для ахроматиновой фигуры (см. ниже).— П р о-должительностьК. измеряется часами (у тритона—1,5—2 часа, у саламандры—2—5 ч.), у теплокровных—короче (х/2 часа); нагревание в известных пределах значительно ускоряет процесс. Хроматиновая фигура К. Ко^ личество, относительные, а также (с меньшей строгостью) и абсолютные размеры, форма и нек-рые индивидуальные особенности хромосом отнюдь не случайны; неизменно повторяясь при всех К. данного вида, они являются характерными и постоянными. Зато даже у близких видов здесь могут иногда наблюдаться значительные различия, а при большей систематической удаленности различия эти обычно резко бросаются в глаза. Так, числа хромосом колеблются у разных видов от двух (Ascaris megalocephala, var.monovalens) до сотен (напр. у рачкаArte-mia salina 168 хромосом); наиболее часто встречаются однако немногие десятки (60 у лошадей, 48 у человека и мартышки, 44 у кролика, 40 у мыши, 24 у лилий, 22—12 у разных видов сумчатых и т. д.). Размеры хромосом также могут значительно разниться: наиболее мелкие являются шариками в 0,3—0,4 fi в диаметре, крупные палочковидные могут достигать в длину нескольких десятков /*. Совокупность хромосом данного вида, или хромосомный комплекс (хромосомная пластинка, кариограмма), является ядерной характеристикой вида, или кариоти-пом (см.). Во всех соматических (телесных) клетках животных и растений [за исключением гаплоидных (см.) генераций споровых] хромосомная пластинка состоит из пар тождественных (гомологичных) хромосом отцовского и материнского происхождения; непарными могут быть лишь половые хромосомы (гетеро-, идио-, аллосомы, х-, у-, w- и г-элементы), к-рые у нек-рых видов значительно отличаются от прочих (аутосомы) и своими размерами. Интересно отметить, что и расположение хромосом в течение К. является не случайным, а постоянным и закономерным, проявляя ясную биполярность; в связи с этим элементы постоянства в расположении могут нередко быть констатированы и в других стадиях К Факт постоянства числа, величины и отличительных особенностей хромосом в гарнитурах служит главным основанием гипотезы индивидуальности хромосом (Бовери), сущность к-рой сводится к тому, что хромосомы продолжают существовать и после реконструкции в покоящихся ядрах в виде отдельных участков ядра, каждый из к-рых при начале К. вновь дает хромосому. В пользу этой гипотезы говорят также опыты скрещивания разновидностей с различным числом хромосом в яйце и спермин: число хромосом потомства всегда равно сумме хромосом материнских и отцовских (аскарида, морские ежи, растительные гибриды); их можно бывает различить и по величине. Наконец прямые указания дает форма нек-рых покоящихся ядер, снабженных пальцевидными выростами (аскарида),—число выростов соответствует числу хромосомных концов; у рачков обособление материнских и отцовских хромосом заметно не только во время дробления, но и в течение всей жизни (гономерия; Наескег). Новые экспериментальные доказательства индивидуальности получены недавно при действии больших доз рентгеновских лучей на хромосомы плодовой мушки Drosophila (Muller, Painter): различного рода дефекты и аномалии, вызванные экспериментальным фактором, оказались стойко передающимися в потомстве. Тонкое строение хромосом несмотря на все усилия исследователей до сих пор не может считаться установленным. По одному взгляду, хромосома образуется и состоит из отдельных зерен—хромомер, или зерен Пфиц-нера (Pfitzner, Balbiani); такой состав ясно выступает на нек-рых фиксированных препаратах (рис. 3), но наблюдался и при жизни (Chambers). Расщепление хромосомной нити сводится тогда к делению хромомер. Нек-рые авторы (Eisen, M. Heidenhain) предполагают еще более мелкие зерна хроматина, хромиолы, из к-рых слагаются хромоме-ры. Сами хромомеры, как предполагают многие, состоят из ахроматиновой основы (линии) и пропитывающего ее хроматина; соответственно этому учение об индивидуальности хромосом базируется на сохранности ахроматиновых участков (Наескег). По другому взгляду, хромосома представляет морфологически цельное образование, состоящее или из чистого хроматина или из линина и хроматина; при этом хроматин или располагается по периферии в виде футляра или разделяется на 2 нити, спирально обвивающиеся вокруг оси (Bonnevie). Другие авторы, наоборот, помещают хроматин по оси, а с поверхности предполагают неокрашивающуюся оболочку в роде футляра (На-вашин) или предполагают, что хромосома сохраняет свою форму благодаря особому скелету в виде нитей (Кольцов). В химич. отношении хромосомы отличаются от хроматина покоящегося ядра большим содержанием базихроматина, богатого фосфором (Lilienfeld, M. Heidenhain), вследствие чего при окраске фуксин-метилгрюном получают более чистый зеленый оттенок. При жизни хромосомы являются однородными, сильно преломляющими свет образованиями, к-рые обладают значительной плотностью и упругостью, при передвижении легко деформируются и могут быть растянуты микроиглами довольно сильно, прежде чем разорвутся. Ахроматиновую фигуру К. изучают обыкновенно в яйцевых клетках, где она достигает большой величины. На фиксированных препаратах она состоит из пузырьковидных центросом с центриолями и отчетливых волоконец, отходящих от них. Часть из них направляется лучистым сиянием к периферии (полярные сияния, астеры), часть образует непрерывное соединение между центросомами (проходящие волокна веретена), часть идет к хромосомам (тянущие нити, плащевые волокна). Первое время им всем приписывали сократительный характер и сравнивали даже с мышечными фибрила-ми (van Beneden, Kalb, Flemming, Boveri); при более детальном изучении выяснилось, что во всяком случае всем волокнам верете-нанельзя приписывать такую функцию—проходящие волокна, наоборот, являются твердыми, упругими образованиями, раздвигающими при своем росте центросомы и препятствующими их сближению (Druner, Meves). Прижизненные исследования вполне подтверждают такое предположение (Chambers, Belar); микродиссекция показывает, что веретено является довольно плотным студенистым образованием. Что касается волокон полярных сияний, то они также обнаруживают значительную твердость, при надавливании иглой сгибаются; если сильно подвигать иглой в клетке, полярные сияния исчезают, но через некоторое время вновь появляются (Chambers). Место, где помещается центросома, имеет жидкую консистенцию. О возникновении веретена шли долгие споры: многие думали, что оно образуется целиком или частью из лининового остова ядра; но в ряде случаев, где удалось точно проследить его возникновение, оно несомненно возникает из протоплазмы, по мнению нек-рых исследователей—именно из особого участка ее, лежащего вблизи ядра (сфера, архоплаз-ма); вопрос этот нуждается еще в дальнейшей разработке. Ядрышко при К. по обычному представлению исчезает после растворения ядерной оболочки и вновь появляется в дочерних ядрах; но в ряде случаев было описано нахождение его обломков в течение всего К.; иногда оно вытягивается по оси веретена и делится. Статистические исследования над растительными клетками показали, что в известном проценте случаев (для разных видов от 15 до 65) ядрышко можно обнаружить в метафазе, а в небольшом проценте части его сохраняются до анафаз и непрерывно переходят в дочерние ядра (Котляревская). Механизм К. Для объяснения движений, наблюдаемых при К. (странствование хромосом,перешнурование тела клетки), в 90-х и 900-х гг. было предложено много гипотез, к-рые в наст, время представляют б. ч. только исторический интерес. Первые исследователи объясняли расхождение хромосом к полюсам и перешнурование клеточного тела сокращением ахроматиновых волоконец. В деталях такая механическая тео- рия была развита Гейденгайном, к-рый построил даже особую модель из полукруглых пружин и резиновых нитей, иллюстрирующую перешнурование клетки. Другой ряд
Рисунок 5. Рисунок 6. Рисунок 7.
Рисунок 5. Схема деления клеточного тела; течение периферической плазмы к экватору. Рисунок 6. Кариокинез растительных клеток-стадий экваториальной пластинки. Рисунок 7. Кариокинез растительных клеток—стадий дочерних клубков—образование клеточной пластинки. теорий физ. характера трактовал ахромати-новую фигуру иначе: в ней видели воплощение силовых линий, а источник силы помещали в центросомах (динамические центры). При этом одни ученые (Н. Е. Ziegler, Gallar-do, Hartog) усматривали аналогию с силовыми линиями электромагнитного поля, которые выявляются напр. железными опилками вокруг магнитных полюсов, раздвинутых на известное расстояние; другие (Biit-schli, Rhumbler) видели в центросоме участок, всасывающий жидкость и вызывающий таким путем диффузионные токи, производящие деформацию в ячеистой плазме (гидродинамические теории). Новейшие авторы (Chambers, Heilbrunn), базируясь на экспериментальных данных, склонны видеть в К. сложный процесс затвердения и разжижения коллоидальной плазмы. Веретено, вырастая и твердея, дает точки опоры для полярных сияний (как предполагал Druner). Волокна полярных сияний не могут тянуть оболочку клетки (как предполагали раньше), так как к периферии разжижаются и весь периферический слой протоплазмы к началу деления становится жидким; в нем наблюдаются токи, идущие от полюсов к экватору; здесь они загибаются соответственно круглой форме астеров и образуют круговую борозду (Speck) (рис. 5). В последних работах, проведенных на живых объектах, Беларж (1929) ставит передвижение хромосом в зависимость от двух факторов: от удлинения среднего отрезка центрального веретена (Stemm-кбгрег) и от сокращения тянущих нитей, происхождение к-рых однако получает совершенно новое толкование. Каждая хромосома, пока она не вошла окончательно в соприкосновение с центральным веретеном, несет в определенной T04Ke(Insertionsstelle)Ka-пельку чрезвычайно вязкой жидкости, к-рая при соприкосновении с волокнами веретена растекается по нему и дает начало сократительному волокну, резко отличному от прочих. Данные эти совпадают с данными Насонова, к-рому удалось у ряда растительных объектов изолированно импрегнировать тянущие нити осмием, и С. Навашина, уже давно описавшего на нек-рых крупных хромосомах повидимому упомянутые выше капли, дающие по Беларжу начало тянущим волокнам («сяжки» Навашина). Особенности К. у высших растений (рис. 6 и 7) сводятся к полному отсутствию центросом и полярных сияний и к делению тела путем образования клеточной пластинки. В экваториальной части бочонко-образного веретена образуется ряд зернышек, которые, сливаясь меяеду собой, дают начало перегородке, сначала простой, затем расслаивающейся на 2 части; перегородка состоит из пеллюлёзы. У некоторых беспозвоночных (яйца нек-рых моллюсков и ми-риапод, гидры) вся кариокинетическая фигура развивается внутри ядерной оболочки, растворяющейся лишь очень поздно или совсем не растворяющейся,—в нутриядер-ный К. Значительные отступления в сторону упрощения свойственны ядрам многих простейших, у к-рых деление протекает в форме промитоза (см.). В тканях различных позвоночных при нормальных и пат. условиях встречается К., не доходящий до конца вследствие растворения ахроматиновой фигуры, в результате чего образуются двуядер-ные клетки—а бортивныйК. (железы, мочевой пузырь). Появление в клетке в начале К. нескольких центров приводит к образованию сложной ахроматиновой фигуры из нескольких веретен и ». неправильному распреде- JI II ^ лению хромосом между ^r s\^ «$V*£ отдельными полюсами — ,У/ £V& мультиполярный, или многополюсный К. а ь (мегакариоциты, клетки РИс. 8. Хромосом-опухолей). Деление и здесь ные пласинки часто не ДОХОДИТ ДО конца. в С1!ерматоцитах: Особое место среди укло- ^ое^Ж нений занимает р е д у к - nus nmetiventris. ционный кариокинез, к-рый входит в цикл созревания половых элементов (рис. 8) и приводит к образованию клеток с половинным (гаплоидным) ЧИСЛОМ ХРОМОСОМ. в. Карпов. Физио югия. Физиол. исследование К. исходит из того факта, что скорость размножения биол. объектов, т. е. число появляющихся в нек-рую единицу времени клеток, непостоянна, а синхронизм (одновременность всех происходящих делений), свойственный первым стадиям дробления яйца, утрачивается довольно скоро (не позже десятого деления). Ритм размножения отдельных клеток меняется, что может зависеть либо от увеличения продолжительности К. либо от увеличения промежутка между двумя последовательными К. Кроме изменения ритма К. отдельных клеток приходится наблюдать общие и местные увеличения числа К. как в результате нормального эмбрионального развития (достижение определенного этапа, связанного напр. с образованием нового органа), так и под действием экстраординарных причин, играющих очевидно роль раздражителей (клеточных потерь, фнкц. гипертрофии, регуляции со стороны организма как целого и т. д.). Так, при экстирпации части печени кролика Понфик (Ponfick) и др. наблюдали большое количество К., нормально отсутствующих в этом органе. Этот пример указывает на необходимость различать для каждой клетки два отдельных понятия: потенциальную способ ностьк делению и •12 35» реальную возможность разделиться. Осуществление второй необходимо требует наличия определенных изменений в плазме клетки, предшествующих реакции ядра, именно—увеличения ее проницаемости и вязкости; только наличие этих условий приводит к профазе К. Наличие потенциальной способности к К. связано с основными свойствами клеточной плазмы и находится в антагонизме с усиленным обменом веществ, ростом и диференцировкой; примером могут служить тонко диференцированные клетки нервной системы, органов чувств, в обычных условиях никогда повидимому не вступающие в кариокинез. По мнению ряда авторов, исследовавших К. с физиол. точки зрения (Гурвич, Wasser-mann), при изучении причин К. следует строго различать 1) факторы, воздействующие на способность, возможность клетки разделиться (Moglichkeitsfaktoren), и 2) факторы, определяющие, «разрешающие» самый акт К. (Verwirklichkeitsfaktoren). Первые в свою очередь распадаются на стимулирующие и тормозящие.—I. Факторы, стимулирующие готовность к К.Кислород стимулирует возникновение К., но стимуляция К. наблюдается иногда и при недостатке О. В опухолях К. может "протекать аноксибиотически (при отсутствии свободного О). Усиленное питание стимулирует К., причем влияние его сказывается по данным обширного исследования Корнфельда (Kornfeld) лишь на 6—7-й день после начала кормления. Голодание также вызывает кратковременную стимуляцию К. Причины этой стимуляции не вполне ясны; повидимому связанная с недостатком пищи усиленная трата резервных веществ в организме приводит к тому, что отдельные клетки временно снабжаются ими в увеличенном количестве. Опыты с культурами тканей показывают необходимость для усиленных делений продуктов расщеплений белков (эмбриональный экстракт); значительную роль играют также ионы, в особенности ион Са, а также повидимому К и Na. Для нормального осуществления К. необходима определенная реакция среды. По данным Рединга и Слоссе (Reding, Slosse) нек-рая щелочность среды создает оптимальные условия для К., повышение же кислотности задерживает наступление К.—Существуют ли специфические вещества , создающие готовность клеток к делению, остается неясным. Многократно подтвержденное наблюдение из области культивирования тканей и простейших—увеличение интенсивности К.в больших клеточных группах—может иметь различное объяснение (см. также Митогенетичвские лучи). Значение t° для К. несомненно, но трудно укладывается в какие-либо общие для всех случаев рамки. Optimum температурного воздействия на К. совпадает с таковым для основных физ.-хим. процессов в протоплазме. Распределение К. в различных частях многоклеточного организма указывает на значение фактора места. Исследования школы Гурвича указывают на связь между подъемом интенсивности К. и последующим формообразованием. Исследования Корнфельда над роговицей саламандр показали, что в парных органах не только общее число митозов, но и распределение их по стадиям в общем совпадает и выражается параллельными кривыми. Здесь имеет место очевидно общий для обоих регулирующий фактор, как думают нек-рые—гормональной природы. Это последнее предположение находит подтверждение в данных Ромейса (Romeis), показавшего резкое увеличение числа К. в конечности головастиков при воздействии гормона щитовидной железы. Накоплен значительный материал, указывающий на стимуляцию К. при воздействии малых доз различных солей и органических веществ. Сильными возбудителями К. являются этиловый и бутиловый спирты, а также растворимые в жирах краски Scharlachrot и Sudan III, введение к-рых в организм вызывает в результате усиленных К. подобные раку разрастания тканей. Механические раздражения (давление) также стимулируют К. Стимулирующим влиянием на К. обладают малые дозы лучистой энергии: красные и инфракрасные лучи, лучи Рентгена и Ra (радия). Стимуляция К. получена при длительном (5—14 час.) воздействии гальванического тока на корешок некоторых растений; в самое последнее время установлено стимулирующее влияние кратковременного воздействия электромагнитных волн на размножение дрожжевых клеток. Многие из только-что перечисленных факторов—органические соединения, гипертонические растворы, ультрафиолетовые лучи, поранения и т. д.—применены были с успехом рядом исследователей для стимуляции развития неоплодотворенного яйца (искусственный партеногенез). II. Тормозящие К. факторы. К числу таковых прежде всего следует отнести нарушения обычного обмена веществ: отсутствие достаточного количества О (для нормальных тканей), воды, солей и питательных веществ. К. прекращается при перегревании, к-рое может сопровождаться физ.-хим. изменениями структуры клетки: вакуолизацией, обособлением различных фаз в протоплазме. Состояние это в известных пределах является обратимым. К тормозящим факторам, нормально присущим организму, должны быть отнесены: 1. Старение клетки, морфологически очень близкое только-что упоминавшимся картинам «перегрева». Сущность процесса клеточного старения мало выяснена; в недавнее время рядом работ школы Ружичка (Ru-zicka) введено представление о гистерезисе— необратимом изменении коллоидов протоплазмы—как причине старения. Гистерезис ведет к постепенному затуханию К.;—2. Ди-ференцировка. По мнению ряда авторов старение клетки связано с образованием стойких продуктов метаморфоза; быть может диференцировку и образование специальных клеточных структур позволительно рассматривать как преждевременное старение и сопоставить с тем, что (как было упомянуто уяда выше) диференцированная клетка обычно не делится, хотя и не теряет еще этой способности окончательно, подобно состарившейся; К. могут появиться в случае возможной дедиферешщровки.—3. Ряд исследований посвящен антагонизму между работой клетки и К. Специальные изыскания Мевеса (Meves) и в особенности Петера (Peter) показали, что при К. специальные функции различных клеток (почечных, железистых и т. д.) значительно понижаются. Во время К. в железистых клетках наблюдается распадение аппарата Гольджи на диктиосомы, обеднение клетки гликогеном, уменьшение содержания НЬ в эритроцитах и т. д. Нек-рые данные Петера показывают наличие и обратной зависимости—работа тормозит течение К. Так, при действии пилокарпина, стимулирующего секрецию, число митозов в железах резко падает.—По данным авторов, работающих с культурами тканей, в серуме нормально присутствует некоторое количество тормозящих К. веществ, по-видимому липоидной природы. Причину клеточного старения и связанного с ним прекращения кариокинеза эти авторы видят в действии указанных веществ, наряду с некоторыми изменяющимися с течением времени протеинами крови. Воздействием различных ядов обычно удается вызвать не прекращение К., а чаще— неправильное их течение. Интересны данные Полицера (Politzer), показавшего тормозящее воздействие «витальной» краски Neutralrot даже в значительном разведении (1 : 150.000); в этом случае наблюдается как извращение морфологии К., так и полное их прекращение; восстановление числа К. происходит через несколько суток, причем вначале число их далеко отстает от нормы. В противоположность малым дозам воздействие лучистой энергии в значительных количествах ведет к торможению К. Эффект этот может быть получен как от видимых лучей (электрическая лампа), так и от лучей Ra и Рентгена; в этом последнем случае эффект напоминает только-что описанное для воздействия Neutralrot (извращение морфологии, исчезновение К. и неправильности в ходе вновь появляющихся). При воздействии лучей Рентгена на семенник установлено тормозящее их влияние лишь на типичные К.; деления созревания оказываются гораздо более стойкими.—Уменьшение числа К. в коже наблюдается у самок морской свинки в период течки. По данным Штиве (Stieve) влияние нагревания всего организма сказывается на уменьшении числа кариокинезов в семеннике. Каков механизм происходящих здесь косвенных воздействий, в настоящее время сказать не представляется возможным; очевидно лишь, что здесь имеет место весьма сложная система гормональных влияний, Факторы осуществления. Только-что рассмотренные данные относились к способности клетки разделиться. Относительно непосредственной «разрешающей» причины К., понимаемой рядом авторов под термином «факторы осуществления», в наст. время существуют 3 теории. 1. Внутриклеточная теория (Kernplasmarela-tion; R. Hertwig) объясняет наступление К. нарушением нормального равновесия между количеством ядерного и плазматического веществ. Теория эта находит подтверждение в ряде фактов, гл. обр. из области эмбриологии. Однако обобщение ее представляется пока за- труднительным; точно так же представление о Kernplasmarelation как единственном факторе, осуществляющем К., требует специального доказательства.—2. Теория гормонов Габерландта (Haberlandt) исходит из ряда установленных на растениях фактов о влиянии на К. возникающих при поранении продуктов распада (раневых гормонов). Габерландт обобщил эти данные и создал теорию, рассматривающую каждый К. как результат воздействия некрогормо-нов. В недавнее время взгляды Габерландта были приняты и обобщены Гутгерцом (Gut-herz), употребляющим для такого делительного гормона термин «метаболии»; при этом он указывает на то, что и в нормальном метаболизме кроются причины возникновения К. В подтверждение своей мысли Гутгерц приводит ряд данных относительно стимулирующего воздействия продуктов аутолиза (парентеральное введение аутолизата, неспецифическая протеинотерапия и т. д.). Однако специфичность гормонов Габерландта как «фактора осуществления» представляется далеко не установленной прежде всего в смысле их (гормонов) универсальности. Иногда при нанесении ран обнаружить К. не удается, и дефект замещается, вначале по крайней мере, за счет клеточных перемещений. Независимо от этого ряд данных заставляет предположить, что гормоны Габерландта должны быть отнесены к группе «факторов готовности», а не «осуществления». — 3. Митогенети-ческие лучи. В 1922 г. Гурвичем было открыто наличие в растительных и животных организмах ультрафиолетовых лучей, длиной около 2.000 А, названных им мито-генетическими и являющихся по его мнению специфическим возбудителем К. За последние годы митогенетические лучи подвергнуты детальному изучению (см. Митогенетические лучи). Универсальность их, доказанная для значительного числа случаев К. животных, растительных и у простейших как работами самого Гурвича и его школы, так и ряда других ученых, делает весьма вероятным их значение как необходимого фактора каждого кариокинеза. Вопрос однако не может в настоящее время считаться окончательно решенным. С. Залкинд. Лит.: Максимов А., Основы гистологии, ч. 1—Учение о клетке, П., 1916 (лит.); Огнев И., Курс нормальной гистологии, ч. 1—Учение о клетке, М., 1908 (лит.); В ё 1 a i К., Beitrage zur Kenntnis des Meehanismus der indirekten Kernteilung, Natur-wissenschaften, B. XV, 1927; он же, Beitrage zur Kausalanalyse der Mitose, 2. Mittcil., Arch. f. Ent-wicklungsmech., B.CXVIII, 1929 и 3. Mittcil., Archiv f. Zellforscliung, Band X, 1929; Flemmlns W., Zellsubstanz, Kern u. Zellteilnng, Lpz., 1882; Gur-witstl A., Das Problem der Zellteihing physiolo-giseh betrachtet, Berlin, 1926; Gutherz S., Der Partialtod in funktioneiler Betrachtung, Jena, 1926; Haberlandt G., Zur Physiologie der Zelltei-Iung, 1—B. Mitteilung, Sitzungsberichte d. Preuss. Ak. d. Wissensch.', rhys. - matheraat. Klasse, 1913— 1914, 1919—21; Hertwig R., tJber Korrelation von Zell- u. Kerngrosse, Biol. Zentralbl., B. XXIII, 190.3; Mac С 1 e n d о n, The laws of surface tension a. their applicability to living cells a. cell-division, Arch. f. Entwicklungsmech., B. XXXVII, 1913; M e v e s Fr., Zellteihing, Erg. d. Anatomie, B. VI, 1897,- P ainter Т., A comparative study of the chromosomes of mammals, Amer. naturalist, v. LIX, 1925; Wassermann P., Wachstum u. Vermeh-rung der lebendigen Masse (Hndb. d. mikroskop. Anatomie des Menschen, herausgegeben v. W- MOllendorff, B. I, p. 441—529, Berlin, 1929, литература). См. также литературу к. ст. Клетка.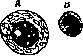
Хроматолиз клеточных ядер: А -семенная кл тка из семонника Sa-lamandra гласи Ь-sa; В — пром шуточное телщ1 из семенника Ascaris megaloc.phalus.
Большая медицинская энциклопедия. 1970.
Карио — Карио, Чеки Чеки Карио Tchéky Karyo Чеки Карио в фильме «Поцелуй дракона» Имя при рождении: Барух Чеки Карё … Википедия
карио... — карио... (гр. karyon орех, ядро ореха) первая составная часть сложных слов, обозначающая: относящийся к клеточному ядру, напр.: кариология, кариоплазма. Новый словарь иностранных слов. by EdwART, , 2009. карио... [< гр. opex, ядро ореха] – в… … Словарь иностранных слов русского языка
КАРИО... — КАРИО... (от греч. karyon орех ядро ореха), часть сложных слов, означающая: относящийся к клеточному ядру … Большой Энциклопедический словарь
Карио... — карио... Начальная часть сложных слов, вносящая значение: имеющий отношение к клеточному ядру, связанный с ним (кариокинез, кариология, кариоплазма и т.п.). Толковый словарь Ефремовой. Т. Ф. Ефремова. 2000 … Современный толковый словарь русского языка Ефремовой
КАРИО — (от греч. karyon орех, ядро ореха), часть сложных слов, указывающая на их отношение к клеточному ядру (напр., кариокинез, кариоплазма). .(Источник: «Биологический энциклопедический словарь.» Гл. ред. М. С. Гиляров; Редкол.: А. А. Бабаев, Г. Г.… … Биологический энциклопедический словарь
карио... — карио... (от греч. káryon орех, ядро ореха), часть сложных слов, означающая: относящийся к клеточному ядру … Энциклопедический словарь
карио- — Имеющий отношение к клеточному ядру: кариология, кариотип. [Арефьев В.А., Лисовенко Л.А. Англо русский толковый словарь генетических терминов 1995 407с.] Тематики генетика EN karyo … Справочник технического переводчика
Карио... — КАРИО... (от греч. káryon – орех, ядро ореха), часть сложных слов, указывающая на их отношение к клеточному ядру (напр., кариокинез, кариоплазма) … Биологический энциклопедический словарь
карио- — karyo карио . Имеющий отношение к клеточному ядру: кариология, кариотип. (Источник: «Англо русский толковый словарь генетических терминов». Арефьев В.А., Лисовенко Л.А., Москва: Изд во ВНИРО, 1995 г.) … Молекулярная биология и генетика. Толковый словарь.
КАРИО — (мексиканский коммунист; в знач. нариц.) Страна (Мексика)! / Поди, / покори ее! / Встают / взамен одного Запаты / Гальваны, / Морено, / Карио. М925 (195) … Собственное имя в русской поэзии XX века: словарь личных имён
КАРИО-
18+
© Академик, 2000-2025
- Обратная связь: Техподдержка, Реклама на сайте
Экспорт словарей на сайты, сделанные на PHP, Joomla, Drupal, WordPress, MODx.